Effect of microbiome on local extraadrenal corticosteroid metabolism in the intestine and secondary lymphoid organs
Glucocorticoids are hormones that are released in response to stressors and exhibit many activities, including immunomodulatory and anti-inflammatory activities. They are primarily synthesized in the adrenal gland but increasing evidence has indicated the existence of extra-adrenal steroidogenesis, which operates in two different modes, the glucocorticoid de novo synthesis from cholesterol and the regeneration of cortisol and corticosterone from their inert 11-oxo metabolites, which is catalyzed by the enzyme 11β-hydroxysteroid dehydrogenase type 1 (11HSD1). Several studies have shown that immune stress increases the capacity of extra-adrenal organs to produce glucocorticoids but the role of microbiota in local synthesis and regeneration of glucocorticoid hormones remains largely unknown. Therefore, the scope of this project is to determine the effect of stress on extraadrenal steroidogenesis and regeneration of corticosterone via 11HSD1.
 |
|
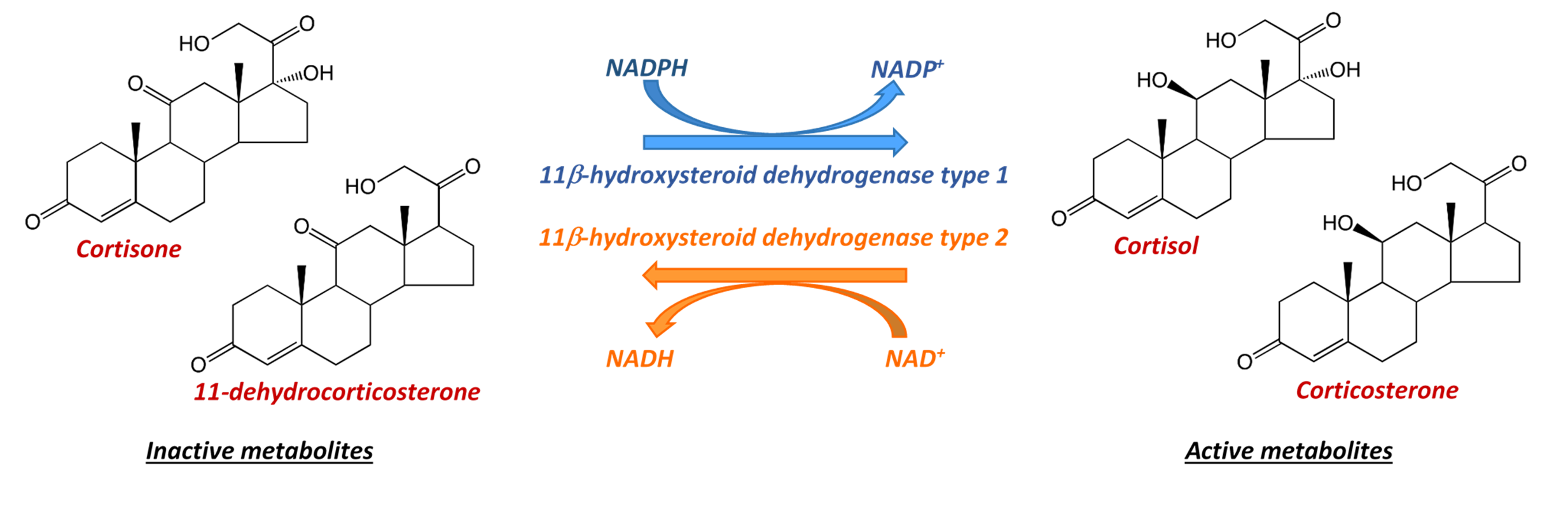
The enzyme 11β hydroxysteroid dehydrogenase type 1 (11HSD1) converts inactive 11-keto metabolites of glucocorticoid hormones (cortisone in man, 11‑dehydrocorticosterone in rat and mice) to biologically active glucocorticoids (cortisol in man, corticosterone in rodents), whereas the enzyme 11HSD2 operates in the opposite direction |
|
|
|
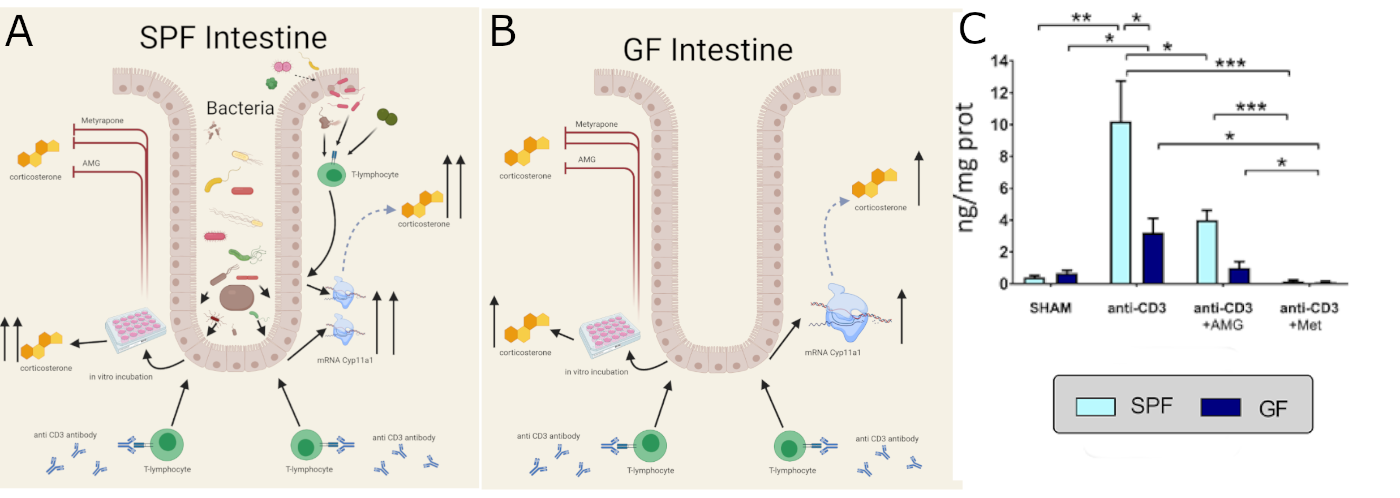
Possible mechanisms of local activation of de-novo corticosterone synthesis in SPF (A) and GF (B) intestine and corticosterone production of in intestine explants from SPF and GF mice stimulated by intraperitoneal injection of anti-CD3 antibody (C). The production of corticosterone is stimulated injection of anti-CD3 and depends on presence of microbiota. Inhibitors of corticosterone synthesis aminoglutethimide (AMG) and metyrapone (Met) significantly decreased corticosterone production in small intestine (Ergang et al. 2021). |
|
|
|
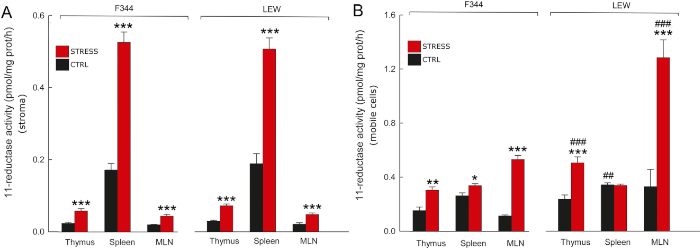
Chronic psychosocial stress increased 11-reductase activity of 11HSD1 (and thus the regeneration of corticosterone) both in stroma (A) and mobile cells of secondary lymphoid organs. The two rat strains differing in stress response were used: the hyper-responsive Fischer 344 (F344) and the hypo-responsive Lewis (LEW) rats (Ergang et al. 2018). |