
Glutamátové receptory
Glutamát je jedním z nejvýznamnějších neuropřenašečů a přibližně polovina všech synapsí v mozku využívá tuto aminokyselinu pro přenos informace. Skutečnost, že dokážete číst tyto řádky, poslouchat hudbu, a hlavně že si dokážete následující den vzpomenout, co jste četli den předchozí, znamená, že váš glutamátový systém je pravděpodobně v pořádku.
Existují však lidé s vážnou poruchou funkce glutamátových receptorů. Ta se může projevit neuropsychiatrickými poruchami, jako jsou halucinace u schizofrenie, demence u Alzheimerovy choroby, po mozkových příhodách či po ischemickém poškození mozku. Proto se snažíme pochopit mechanizmy, které jsou v mozku zodpovědné za správnou funkci glutamátového systému.
Glutamát působí na membránových proteinech nazývaných glutamátové receptory. Předmětem našeho studia je jedna ze skupin těchto receptorů, které pokud jsou aktivovány glutamátem, umožňují iontům vstupovat do buněk. Proto je tato skupina označována jako ionotropní glutamátové receptory. Tok iontů zprostředkuje přenos informace, protože "říká" buňce, jak se zachovat a jak přizpůsobit svoji aktivitu, a to i na relativně dlouhou dobu. Tyto dlouhodobé změny nervové aktivity jsou považovány za základ pro tvorbu a uchování paměťových stop v mozku.
Ionotropní glutamátové receptory, které studujeme, se podle způsobu řízení, aktivace a struktury dělí do tří skupin. Struktura glutamátových receptorů je velmi důležitá, poněvadž určuje ostatní charakteristiky receptoru. Podle specifické struktury jsou jednotlivé podtypy receptorů aktivovány konkrétními látkami, podle kterých jsou příslušné receptory pojmenovány: NMDA, AMPA a kainátové receptory.
Předmětem našeho studia jsou primárně vlastnosti NMDA receptorů. V posledních letech se naše znalosti o podjednotkovém složení těchto receptorů velmi zdokonalily, takže dnes můžeme říct, jaký konkrétní typ NMDA receptoru se v daném neuronu nachází. Je známo, že každý funkční NMDA receptor se skládá ze čtyř podjednotek (Obr. 1). Tyto tzv. nativní receptory však nejsou ve zdravých buňkách lokalizovány samostatně. Existuje totiž mnoho dalších proteinů, které se s NMDA receptory přímo spojují. Pro mnohé z těchto intracelulárních proteinů však zatím není přesně známo, jak ovlivňují aktivitu receptorů.
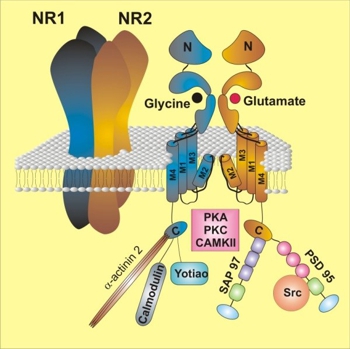
Obr. 1 Struktura a podjednotkové složení NMDA receptorů. NMDA receptory se skládají ze čtyř podjednotek (tetramerní uspořádání) – obvykle sestaveny ze dvou kopií NR1 a dvou kopií NR2 nebo NR3 podjednotek. Každá podjednotka NMDA receptoru obsahuje extracelulární N-terminální doménu (N-konec), čtyři membránové domény a intracelulární C-terminální doménu (C-konec). Část N-konce, která vystupuje nejdále od membrány a obsahuje vazebná místa pro některé modulační látky, jako např. protony, polyaminy (spermin, spermidin), neurosteroidy, Zn2+, Cu2+ atd. Vazebná místa pro agonistu (glutamát) a koagonistu (glycin) jsou lokalizována ve střední části N-konce podjednotek NR2, resp. NR1. Transmembránová část receptoru se skládá ze tří domén (M1, M3, M4), které zcela procházejí membránou a domény M2, která do membrány vstupuje na intracelulární straně, vytvoří kličku a opět membránu na intracelulární straně opouští. M2 kompletního tetrameru receptoru tvoří pór iontového kanálu. Dlouhý C-konec NMDA receptoru slouží jako doména pro interakci s intracelulárními proteiny, včetně signálních molekul a proteinů, podílejících se na transportu receptorů z intracelulárních míst jejich syntézy do plazmatické membrány, jejich fosforylaci či defosforylaci. Na obrázku jsou znázorněny interakce podjednotek NR1 a NR2 s některými z těchto proteinů. Pro zjednodušení zobrazuje pravá část obrázku pouze jednu podjednotku NR1 a jednu podjednotku NR2.
Pro studium NMDA receptorů s konkrétním podjednotkovým složením využíváme buňky, které za normálních okolností NMDA receptory nemají, jako např. lidské embryonální ledvinné (HEK) buňky. Pomocí metody transfekce vkládáme do HEK buněk geny pro příslušné podjednotky NMDA receptorů, které se v membránách HEK buněk následně exprimují. Za těchto experimentálních podmínek můžeme studovat NMDA receptory s přesně definovaným podjednotkovým složením.
Velmi důležité pro tuto práci je, že do HEK buněk nemusíme vkládat geny pro běžně se vyskytující, nativní NMDA receptory, ale že můžeme měnit strukturu genů a studovat, jak se mění funkce následně vzniklého a exprimovaného NMDA receptoru. Můžeme např. použít část podjednotky NR2A, část podjednotky NR2C, spojit je k sobě a vytvořit tzv. chimérní NMDA receptor (chiméra - odvozeno z mytologie podle stvoření, které bylo zčásti lev, zčásti člověk a zčásti drak). Jiným metodickým přístupem můžeme změnit jednu aminokyselinu, které kódují protein některé z podjednotek a studovat, jak významná tato změna bude pro konkrétní aspekt aktivity a funkce receptoru (tzv. bodové mutace).
Neurosteroidy jako modulátory NMDA receptorů Obecně je dobře známá skutečnost, že některé hormony v těle mají steroidní strukturu. Notoricky známé se staly steroidní hormony díky dopingovým aférám, spojeným s jejich nelegálním užíváním ve sportu. Avšak naše mozky také produkují látky steroidní povahy, které mohou ovlivňovat aktivitu některých receptorů, včetně NMDA receptorů. V takových případech se tyto látky nazývají neurosteroidy. V současné době, ve spolupráci s dr. Hanou Chodounskou a jejími kolegy (Ústav organické chemie a biochemie AV ČR, v.v.i.), testujeme působení dvou endogenních steroidních látek s podobnou chemickou strukturou, ale s naprosto odlišným modulačním působením na aktivitu NMDA receptorů. Jedna z těchto látek se nazývá pregnenolon sulfát (PS), druhá pak pregnanolon sulfát (3α5βS; tuto zkratku používáme k označení pozic 3 a 5 pro atom vodíku ve struktuře steroidního kruhu) (Obr. 2, struktura steroidů). Pro testování vlivu těchto neurosteroidů na NMDA receptory využíváme metodiku patch-clamp. Pro aplikaci agonisty a ostatních látek na buňky používáme rychlý aplikační systém.
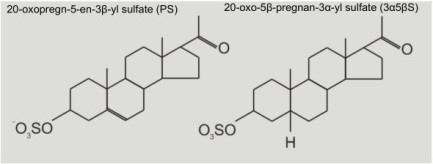
Obr. 2 Struktura pregnenolon sulfátu (PS) a 20-oxo-5-beta-pregnan-3-alpha-yl sulfátu (3α5βS).
V naší experimentální práci jsme zjistili, že pro oba studované neurosteroidy platí, že jejich konečné působení na NMDA receptorech je dáno způsobem jejich aplikace ve vztahu k aplikaci glutamátu jako endogenního agonisty NMDA receptorů. Také jsme ukázali, že míra působení těchto neurosteroidů přímo závisí na podjednotkovém složení NMDA receptorů.
Pro působení PS jsme ukázali, že pokud je aplikován v nepřítomnosti agonisty (glutamátu nebo NMDA), jsou následné odpovědi vyvolané aktivací NMDA receptorů výrazně potencovány. Pokud je PS aplikován současně s agonistou, způsobuje potenciaci receptorů, které obsahují podjednotku NR2A nebo NR2B, ale inhibuje aktivitu receptorů, obsahujících podjednotku NR2C nebo NR2D (Obr. 3). Jak je patrné na obrázku, aplikace samotného glutamátu způsobuje kontrolní proudovou odpověď NMDA receptorů. Proudová hladina je <0, což znamená, že přes iontový kanál spojený s NMDA receptorem pronikají do buňky ionty s kladným nábojem. Následně, je-li PS preaplikován, amplituda proudové odpovědi se zvětší. Přitom platí, že čím vyšší koncentrace PS, tím vyšší míra potenciace nastává. Nejvyšší koncentrace PS, kterou jsme experimentálně používali (300 mM), způsobila 4-5 -násobnou potenciaci proudové odpovědi ve srovnání s kontrolou.
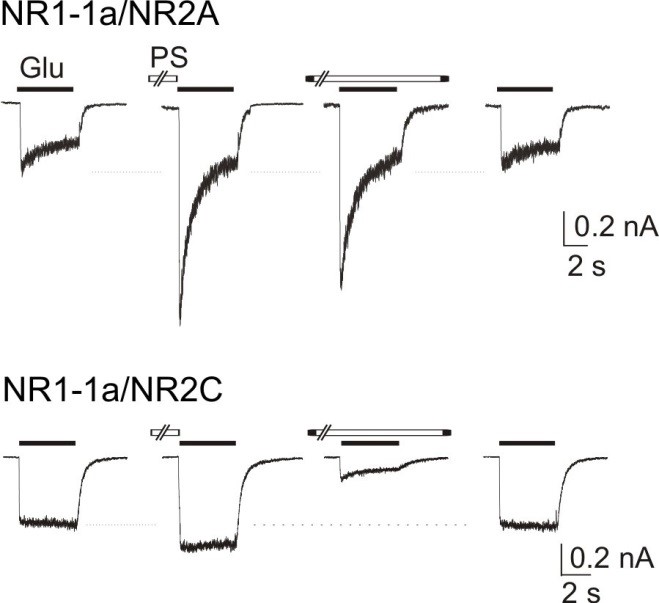
Obr. 3 Působení PS na NMDA receptory obsahující podjednotky NR1/NR2A (nahoře) a podjednotky NR1/NR2C (dole). Preaplikace PS či jeho preaplikace a současná aplikace způsobuje výraznou potenciaci proudových odpovědí NMDA receptorů s podjednotkou NR2A ve srovnání s kontrolní odpovědí. Při preaplikaci na NMDA receptory s podjednotkou NR2C způsobuje PS nepatrnou potenciaci, při preaplikaci a současné aplikaci s agonistou pak výraznou inhibici proudových odpovědí.
V případě 3α5βS jsme zjistili, že NMDA receptory inhibuje, bez ohledu na jejich podjednotkové složení, ale pouze tehdy, je-li aplikován současně s agonistou, jak je znázorněno na Obr. 4. Výsledky našich pokusů ukazují, že 3α5βS se na NMDA receptor nemůže vázat, je-li aplikován v nepřítomnosti agonisty. To je zvláště zajímavá vlastnost, kterou 3α5βS sdílí se skupinou látek zvaných blokátory otevřeného iontového kanálu. Jak již bylo uvedeno, NMDA receptory jsou fyzicky spojeny s kanálem, kterým proudí ionty dovnitř buňky. V nepřítomnosti agonisty je receptor inaktivní a iontový kanál s ním spojený je zavřený. Existují látky, které mohou do iontového kanálu vstoupit a "ucpat" jej. Tímto mechanismem brání toku iontů přes iontový kanál a tak inhibují aktivitu NMDA receptorů. Poněvadž vazebné místo pro určité látky je uvnitř kanálu, nemohou tyto látky manifestovat své působení, aniž by byl receptor aktivován a iontový kanál otevřen. To znamená, že působení takových látek závisí na tom, zda je NMDA receptor "využit" agonistou či nikoli. Z tohoto důvodu jsou uvedené látky nazývány "use-dependentní" inhibitory nebo blokátory otevřeného iontového kanálu.
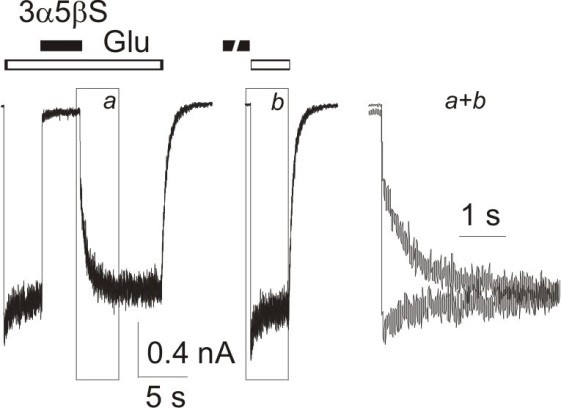
Obr. 4 3α5βS působí na NMDA receptorech jako tzv. "use-dependentní" inhibitor. Na obrázku je znázorněno působení 3α5βS na rekombinantní NMDA receptory s podjednotkovým složením NR1/NR2B exprimované v HEK buňkách. Odpověď na společnou aplikaci 300 mM 3α5βS a 1 mM glutamátu (Glu) provedená po kontrolní aplikaci samotného glutamátu byla téměř úplně inhibována. Odmytí 3α5βS z receptoru a návrat na kontrolní proudovou hladinu je pomalý (na obrázku znázorněno písmenem"a"). Nástup odpovědi po aplikaci 1 mM glutamátu bezprostředně po preaplikaci 300 mM 3α5βS je velmi rychlá, srovnatelná s rychlostí nástupu kontrolní odpovědi na glutamát (na obrázku znázorněno písmenem "b". Zcela vpravo je na obrázku znázorněno překrytí částí "a" a "b", aby byl patrný rozdíl v časovém průběhu odpovědí na glutamát provedenou po současné aplikaci 3α5βS a glutamátu a na aplikaci glutamátu provedenou po preaplikaci 3α5βS.
Naše pokusy však naznačují, že vazebné místo pro 3α5βS není uvnitř iontového kanálu, poněvadž jeho působení není závislé na membránovém potenciálu, které je naopak obecnou vlastností typických blokátorů otevřeného iontového kanálu.
Testovali jsme také působení 3α5βS na jednotlivé iontové kanály NMDA receptorů. Obecně, jakákoli látka může změnit některou ze tří charakteristik jednotlivých iontových kanálů (nebo jejich kombinace): frekvence otevírání iontového kanálu, vodivost kanálu a dobu, kdy je kanál otevřen (tzv. doba otevření). Ukázali jsme, že 3α5βS inhibuje celkovou aktivitu NMDA receptoru primárně snížením frekvence otevírání iontových kanálů s nimi spojených.